In the early 18th century, the Italian naturalist Lazzaro Spallanzani was studying the sensing of bats. He found out that bats can orientate themselves in absolute darkness without their eyes, but that this is not possible without the help of their ears. But it was only later in the end of the 1930s that the US researcher Donald Griffin and some of his colleagues succeeded in making the calls of bats audible and revealing the bats' acoustic mode of orientation. Echolocation is found not only in bats but also in toothed whales, some cave-dwelling bird species and shrews. Put simply, in echolocation, the brain forms a picture of the environment by analysing echoes produced by self-produced sounds. In most bats, echolocation calls are produced in the larynx. Lips and skin appendages at the nose and mouth area serve as so-called acoustic lenses, which help to focus the calls more precisely, similar to a camera. Bats produce very different echolocation calls that vary in volume, length and pitch depending on the species and situation. Most of the time, these calls are so high-pitched that we, as humans, cannot hear them, which is why they are also called ultrasonic calls. The human hearing range extends from about 2 to 20 kHz. Bat calls are usually higher than 20 kHz and in exceptional cases can reach frequencies of up to 300 kHz. Echolocation calls are also subject to some physical limitations. High frequencies are very quickly attenuated in the air. This means that the higher the frequency used by the animals, the shorter the range of the corresponding call. However, the use of high frequencies also brings advantages. The higher the tone, the smaller structures the bats can acoustically detect. Thus, it is not possible for the animals to achieve a large range with their calls at the same time as high resolution. This problem is solved by the animals in a very flexible way. Most bats can change the calls they emit to adapt them flexibly to the respective environment and situation. In habitats with dense vegetation, such as forests, high-frequency calls are advantageous, because here a precise resolution ensures that obstacles can be avoided and prey can still be detected against the background of the vegetation.
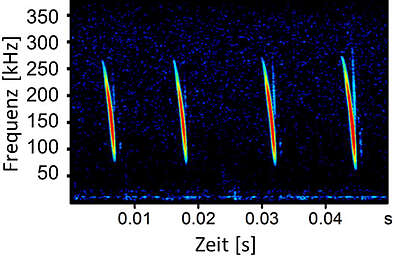
Basically, two types of echolocation calls can be distinguished: Constant-frequency calls and frequency-modulated calls. The latter start at very high frequencies and then pass through a frequency band of lower frequencies. This frequency band can be more or less steep and the modulation of frequencies often occurs in a very short time (2-5 ms). So-called simple-nosed bats (vesper bats), which include most bat species found in Germany, can produce echolocation calls in particularly wide frequency bands. By increasing the bandwidth of the calls, the bats increase the information content of the echo. Especially the distance to an object or a prey insect can be estimated very well this way. Very broadband calls therefore occur mainly in bats that hunt in dense vegetation, such as the native Bechstein's or Natterer's bats (Myotis bechsteinii; Myotis nattereri).
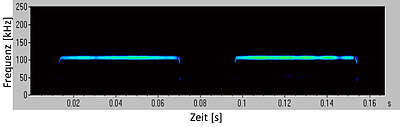
As a mixed type of these two calls, another call type can be distinguished: the quasi-constant-frequency call. Such calls begin with a small frequency-modulated section that can vary greatly from call to call, even in individual animals. The information that can be obtained with this call is not aimed at the exact localisation of an object, but at the detection of objects in a large range. Therefore, this call type is mostly used by bat species that hunt in open areas and fly very fast, such as common noctules (Nyctalus noctula). Then, as soon as an object is detected, the bats switch to frequency-modulated echolocation, which provides more information about the exact location, size or shape of an object.
Fruit bats are the only group within bats that have not evolved echolocation, in which calls are produced via the larynx. However, some fruit bats (of the genus Rousettus) produce echolocation calls via rapid tongue clicks. Therefore, it is possible for these flying foxes to fly in caves where there is total darkness. Flying foxes that cannot echolocate, on the other hand, can only use entrance areas of caves where sufficient daylight makes visual orientation possible.
When listening to echolocation calls with the Batdetector, we distinguish between "wet" and "dry" calls depending on the auditory impression. It is one of the most important distinguishing features for species recognition with the Batdetector.
Pure frequency-modulated calls produce a "dry" auditory impression because the bats call so quickly from top to bottom (i.e. from high to low frequencies) that our ear does not manage to recognise the calls "translated" into our auditory range with the Batdetector as sounds. It simply happens too fast for us. Therefore, these calls have no sound quality for us. These calls are called "dry".
In the case of so-called quasi-constant-frequency calls, the calls are lowered quickly at the beginning but change only very slowly at the end, which is why these calls are almost constant-frequency at the end. The auditory impression is reminiscent of a splash or drops of water falling on a water surface, which is why we speak of "wet" calls here.
When a sound source moves towards us or we move towards the sound source, the frequency of the perceived sound shifts to a higher range than when the sound source moves away from us or we move away from it. This means that we hear a higher tone when the sound source is approaching us (compare approaching ambulance).
you're not there yet? Then register directly:
register for projects
register as teacher
Sie sind noch nicht dabei? Dann direkt anmelden. Jetzt Registrieren